The Origin of Life, Evolution, Metamorphosis.
The universe is alive and forever recycles and gives birth to planets, galaxies and stars.
Giant stars incredibly vast in size, explode, giving birth to hundreds even thousands of infant stars, which come to be ringed with living planets, many just like our own.
And this is how our own story begins....
The seeds of life, actual living creatures and their DNA, flow throughout the cosmos and have taken root on innumerable worlds much older than our own.
And these genetic seeds contained in the DNA instruction for the metamorphosis of all life, including woman and man...and this is how life on our planet began.
Extinction is how our story may end.
For thousands of years humans have gazed into the heavens pondering the nature of existence, asking: "How did life begin?" "Are there people on other planets?" "Are we alone in the vastness of the cosmos?" Humans have long stared into the abyss and the abyss has stared back.
Answers and explanations have ranged from religious beliefs in a creator god, to the magical thinking of modern day scientists who embrace a theology of miracles: preaching that life came from non-life in a supernatural organic soup.
The likelihood that life and its DNA emerged from an organic soup, or
undersea thermal vent -at least on Earth- is the equivalent of discovering a computer on
Jupiter and then arguing that it was randomly assembled in the Methane
Sea.
Only life can give rise to life. Only DNA can give rise to DNA--the machinery of life. Every attempt to prove otherwise has miserably failed.
If life were to suddenly appear on a lifeless, desert island, we would not pretend it was randomly assembled in an organic soup, or created by the hand of god, but that it washed to shore or fell from the sky.
The Earth too, is an island, orbiting in a sea of space, and living creatures, and their DNA, have been washing to shore and falling from the sky, since the Earth's creation.
The first creatures on earth, came from other planets.
For 800 million years after our planet's creation, the Earth was continually bombarded by gigantic meteors, asteroids, and mountains of frozen ice, with the first evidence of Earthly life, highly complex living creatures, appearing immediately thereafter. As only life can produce life, then the first creatures to appear on Earth must have been contained in that debris, and came from other planets.
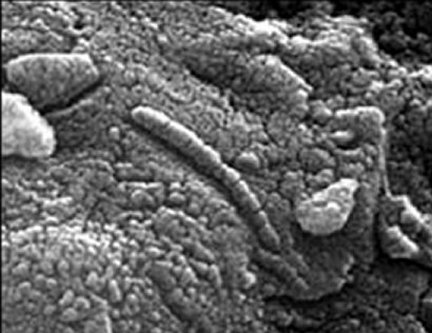
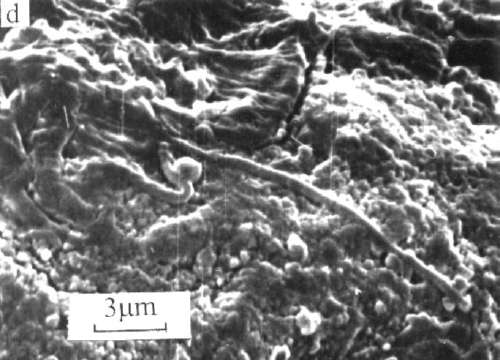
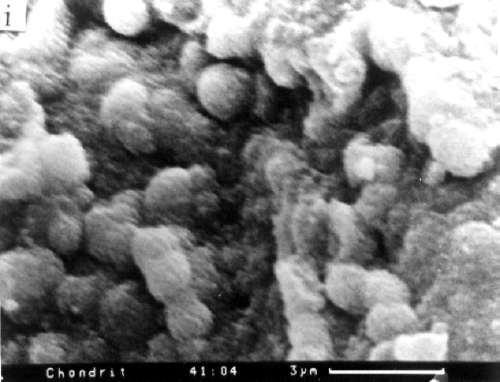
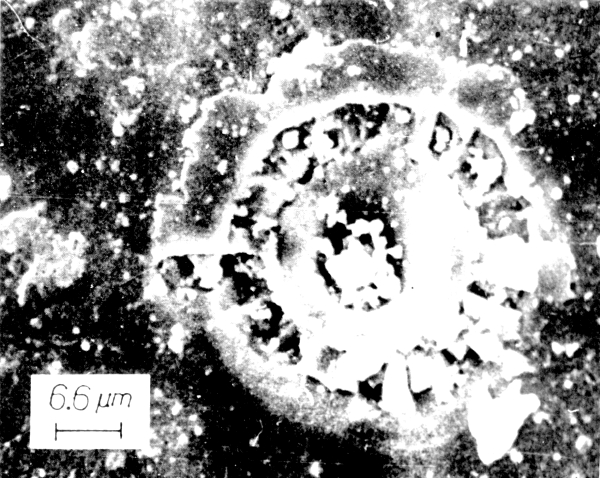
The evidence supporting this rather astounding proposition is extensive, and includes fossils of past life found in 1) three meteors from Mars, 2) five meteors which originated outside the solar system, and 3) three soil samples from the moon. In 1969, when a camera from the Surveyor 3 was retrieved from the lunar surface and returned to Earth, it was found to be coated with "organic material of unknown origin" and a single dormant microbe was discovered inside.
Life can exist in almost any environment, from the freezing to boiling, flourishing at the bottom of the frigid Antarctic ocean, or in liquid fire under 9,0000 crushing pounds of ocean pressure. When threatened with death, life becomes dormant, and may awaken even after 250 million years have passed. Since life exists everywhere on Earth, it is reasonable to assume life can exist anywhere in the cosmos.
Life on Earth came from other planets.
Cosmic collisions are commonplace, not only between meteors
and planets, but entire galaxies, and life has been repeatedly tossed into the abyss...only to land
on other planets.
The genetic seeds of life swarm throughout the cosmos, and these genetic "seeds," these living creatures, fell to Earth, encased in stellar debris which pounded the planet for 700 millions years after the creation. And just as DNA contains the genetic instructions for the creation of an embryo, neonate, child, and adult, and just as modern day microbes contain "human genes" which have contributed to the evolution of the human genome, these "seeds," these living creatures, contained the DNA-instructions for the metamorphosis of all life, including
woman and man. DNA acts to purposefully modify the environment, which acts on gene selection, so as to fulfill specific genetic goals: the dispersal and activation of silent DNA and the replication of life forms that long ago lived on other planets. *****
SCIENTIFIC REVOLUTIONS
The universe is ageless, infinite, and has no beginning and no end. However, the nature of the human mind require endings and beginnings, and thus charlatans and priests invent myths such as the big bang, the organic soup, and Darwinism to give the children "once-upon-a-time" myths to believe in, myths that have beginning and endings.
Throughout the ages, and as is true today, some of what has passed for "scientific fact" has been based on faith and dogma; which is why the temple priests of science often protect the faith, and the status quo, by attacking and ridiculing those heretics who threaten to topple and shatter the altars of their idols. The dustbins of history are laden with discarded "scientific facts" and those who believed in them(Kuhn, 1970). Until the 16th century, it was a "fact" that the Earth was at the center of the solar system and the universe. Until the 19th century it was scientific "fact" that "rocks do not fall from the sky" and that meteors did not exist. Until the 20th century, it was scientific "fact" that interstellar space was permeated by a viscous "ether." In the 1920s, articles and editorials appeared in leading scientific journals ridiculing those rocketeers who dreamed of soaring through space, explaining that it was a scientific "fact " that rockets would be unable to propel themselves beyond the Earth because of the lack of atmosphere or air. Until the year 2000, it was a scientific fact that the speed of light was a constant and that nothing could travel faster than the speed of light.
And all these "scientific facts" have been proved false.
Now we are told that life emerged from an organic soup, and then evolved following the natural selection of "random mutations." And yet, the proverbial organic "alphabet" soup was missing all essential ingredients, including DNA. There was no organic soup, at least not on Earth. The theory of the organic soup is a myth, based on a theology of miracles. There is absolutely no evidence in support of this theory. Rather, this myth has been repeatedly disproved. And yet, the Temple priests of science continue to bow down and worship at this altar, which is little more than a religion masquerading as science. By contrast, there is evidence of past life on 3 meteors from Mars, 5 meteors which originated outside the solar system, and evidence of fossils and life recovered from the moon.
The Organic soup is a myth. Life on Earth originated on other planets.
Nor has life "evolved" randomly, but in a highly predictable, molecular-clock like fashion. There are in fact, genes (e.g., tim, mTim, hTIM) and proteins which perform specific "clock-like" timing functions and which interact to form regulatory feedback loops, and these are highly regulated (Clayton et al., 2001). There is nothing random about the organization or expression of DNA (Caron et al., 2001; Courseaux & Nahon, 2001), the source of all variation.
The history of science is a history of scientific revolutions, where established, authoritative scientific dogma finally crumbles from the weight of unwieldy, disconfirming evidence that can no longer be suppressed or ignored and which continues to grow until it completely undermines the beliefs and the authority of the ruling status quo. The history of scientific revolutions always entails a complete paradigm shift in scientific thought and belief; ushered in by those revolutionaries who dared to challenge the ruling authorities and the high priests of science.
The next revolution has begun...
******
The central tenants of Dr Rhawn Joseph's DNA-based, astrobiological theory of the origin and metamorphosis of life can be summarized as follows:
1) Life is an intrinsic feature of the living, infinite universe.
2) Life arrived on Earth from other planets
3) DNA is capable of learning, remembering, and acting intelligently.
4) The "Seeds of Life" actual living creatures, and their DNA, swarm throughout the cosmos.
5) Cosmic collisions are commonplace, not only between meteors and planets, but between entire galaxies.
6) Living creatures contained in planetary debris have been
repeatedly hurtled to other worlds.
7) Our sun and solar system are the remnants of a vast star system which exploded in a huge supernova at over 5 billion years ago. Debris from the shattered remains of this star system gave birth to many new stars including our sun, the Earth, and solar system. The Earth was bombarded with debris from the ancient star for 700 million years with the first evidence of life appearing on the Earth and on Mars immediately thereafter.
8) Some of the life from this ancient star system survived encased in mountainous meteors and ocean of ice, and were flung upon the surface of the new Earth.
9) The first creatures on Earth..(and their DNA), came from other planets.
10) DNA acts on itself and modifies and alters the environment.
11) The modified environment acts on gene selection to activate "silent" genes and "silent" genetic traits which exist
a priori.
12) These first creatures on Earth, and their DNA then labored to alter the environment so as to engineer their own
evolution.
13) Once the environment is sufficiently engineered, these silent genes and the traits they code for may be expressed
in distinct and separate species.
14) As these "silent" genes/ traits are inherited and were passed down from ancestral species, then these genes and
traits must have been inherited from creatures that "evolved" on other planets.
15) Genes can also be transferred laterally and horizontally between the same and different species, so that different species can come to
possess the same gene and the same trait.
16) Genetic evidence indicates that evolution has progressed in a highly predictable "molecular clock-like" fashion.
17) The progressive "evolution" of increasingly complex and intelligent species in a step-wise progressive fashion,
and genetic evidence as reported by the human genome project, indicates that "evolution" has unfolded in
accordance with specific and highly regulated genetic instructions.
18) Conclusion: Life on Earth, and its DNA, originated on other planets. DNA acts to modify the environment to engineer its own evolution and the activation of traits and
genes which exist a priori; i.e. "evolutionary metamorphosis."
|